「新機制哲學」(the New Mechanical Philosophy) 興起於二十世紀末與二十一世紀初,這個名稱來自生物學哲學家史奇伯 (Robert A. Jr Skipper) 和彌爾斯坦 (Robert L. Millstein),她們把「為了理解機制本質以及它在科學扮演的角色所發展出來的哲學架構」稱為「新機制哲學」(Skipper and Millstein 2005),有時簡稱為「新機制論」(New Mechanism),而那些投入新機制哲學的哲學家就被稱為「新機制論者」(New Mechanists) (Bogen 2008; Craver and Tabery 2015)。新機制論者主要著眼於生物學家習慣使用「機制」這個概念來說明生物現象的事實,深入探討「機制是什麼」,也同時問:生物學家為什麼不用物理化學定律來說明現象而要用機制概念呢?近二十年來,新機制論提供生物哲學一種有別於十六、十七世紀的機械哲學,以及十八、十九世紀神秘的生機論與徹底化約的唯物論,以清晰、易懂的新機制概念來重新理解生物學。
新機制論大約是從 1990 年開始醞釀,當時有三份蠻有關鍵影響力的哲學著作:第一是貝克托 (William Bechtel) 和理查森 (Robert C. Richardson) 於 1993 年出版的《發現複雜:用解組和定位作為科學研究的策略》(Discovering Complexity: Decomposition and Localization as Strategies in Scientific Research),此書最先指出,大部分的生物學家是透過拆解複雜的生物系統,將生物整體解組為各個局部,然後把局部定位至正確的機制結構,以此方式來理解生命現象。第二是格林蘭 (Stuart Glennan),他於 1996 年出版〈機制和因果的本質〉(Mechanisms and the Nature of Causation) 論文,認為英國早期的經驗論者休姆 (David Hume) 所認為因果之間的神秘連結其實就是機制。第三是由麥契默 (Peter Machamer)、達登 (Lindley Darden)、克瑞弗 (Carl F. Craver) 三位哲學家於 2000 年合作的論文〈思考機制〉(Thinking about Mechanisms) 一文。此文不僅為「機制」概念提出簡潔有力的哲學刻畫,後來更發展出一系列完整的發現機制論述,成為後來著名的 "MDC mechanism"(以三位作者的姓名字母聯名)。
什麼是「機制」?「機制」這個概念可以追溯到十六、十七世紀的機械哲學 (the mechanical philosophy),當時自然哲學家如伽利略 (Galileo)、笛卡兒 (René Descartes) 與波以爾 (Robert Boyle) 延續自希臘哲學原子論 (atomism) 的傳統想法,將大自然想像成一個巨大的機器,機器裡有各種零件,零件彼此透過接觸活動 (contact action) 產生各種自然現象,如天體運行、熱汽傳導、與生命現象等。然而,科學發展經過科學革命後,物理學家開始為這些機械運作建立不同的物理概念來理解之(如「吸引力」或「磁力」),並且結合數學發展出一套以定律為核心的近代物理學,使原來舊有的機械哲學慢慢退出物理學的舞台。
不過,雖然機械哲學沒有主導現代物理學,但打從十七世紀到十九世紀的生物學家卻始終沒有放棄以「機制」這個概念來理解生命現象。有些生物學家會用「機械般的過程」(machine-like processes) 來理解生命現象,他們習慣將生命現象拆解為細部成分,然後研究細部成分間的位置與互動,基本上他們認為,生物細部成分間的互動就像機械的局部互動一樣,都可以用物理化學定律來理解,類似這樣的立場屬於「機制唯物論」(mechanical materialism)。但也有另一派的生物學家傾向用「生機論」(vitalism)、「有機論」(organicism) 或「整體論」(holism) 的想法來理解生命現象。基本上這派生物學家的想法是認為,生命現象不能只用機械式的方式來理解之,畢竟生命現象不是拆卸後的零件再加總起來的結果。生命現象裡必定有某種非物質、不可測量的元素或力存在於其中,才使得生命過程是如此複雜地呈現在我們的感官之前。
歷史上,介於機制論與生機論的爭論一直從十七世紀中期到二十世紀都沒有停過,兩派理論就像鐘擺一樣兩邊擺盪,時而前者興盛,時而後者流行。我們無法在此完整交代兩派理論間的辯論與演化,不過隨著美國生理學家洛布 (Jacques Loeb) 與德國胚胎學家斯佩曼 (Hans Spemann) 研究成果所帶來的影響,生機論大概在1880年後期就開始慢慢凋零,而生物學整體的發展便開始穩定地朝向機制論與化約論的方向。儘管機制論與化約論彼此還是有一些細部的差異,如化約意涵很強的「機制唯物論 (mechanistic materialism)」或較重視生物作為一個整體的「整體唯物論 (holistic materialism)」,無論如何,二十世紀以後的生物學都因為基因概念的分子化以及生物技術的進步,確實走向機制與化約論式的研究方向,也因此,生物哲學把二十世紀以後的機制學派稱為「新」機制哲學,目的正在於要與十六、十七世紀的機械哲學區別開來 (Allen 2018)。[1]
接下來我們要開始說明:什麼是「機制」概念?為何「機制」概念對生物學實作如此重要?為什麼生物學家不用物理化學定律來說明生物現象?用機制模型所發展的機制說明與傳統邏輯經驗論的涵蓋律說明有何不同?其中,機制與模型的關係又是為何?第三節我們會討論,新機制論者如何以機制整合的新觀點重新看待生物學的理論關係?第四節介紹新機制論建議的發現策略,包括概念上與實作上的發現推理。最後第五節,我們將介紹台灣科學哲學界近年來與國際新機制哲學家進行的學術交流及其具體學術成果,還有目前國內幾位年輕新機制論者的研究現況。
不同的機制概念會對現象形成不同的機制說明,我們在此介紹目前最常被引用的三種機制概念,並用簡單的例子呈現它們各自的特色。然後,我們會說明由機制所形成的機制說明跟傳統經驗論的涵蓋律說明有何差異,並且指出為什麼機制說明更適合生物學,以及它具備哪些知識論上與說明力的優勢。最後,我們會說明機制模型在機制說明與機制發現裡所扮演的角色。
最早為「機制」提出刻畫的哲學家是貝克托和理查森,他們採取拆解的進路把機制定徵 (characterize) [2]為「由相關的局部 (parts) 組成,每個局部都有各自的功能,彼此用一種能對系統行為產生貢獻的方式結合在一起。」(Bechtel and Richardson 1993: 17) 但「局部」具體指的是什麼?要怎樣「結合」在一起才能產生系統行為?這些問題,貝克托與理查森並沒有說得很清楚,後來的新機制論者紛紛提出不同的機制概念。
目前最常被引用的三種機制概念是:
I. 格林蘭: 「機制是產生行為的複雜系統,由許多局部依據直接、不變、與變化有關的通則互動所產生。」(Glennan 2002: S344)
II. 麥契默、達登、克瑞弗:「機制是由實體與活動組織而成,在起始和終止條件之間,產生規律變化。」(Machamer, Darden, and Craver 2000: 3)
III. 貝克托和亞柏漢森 (Adele Abrahamsen):「機制是一個結構,透過組成局部、運作和組織發揮功能。運作協調的機制負責一個或多個現象。」(Bechtel and Abrahamsen 2005: 423)
下圖是常見的機制示意圖,圖的上層是機制產生的現象,下層是機制。機制內部有各種不同的「實體」(entity)(即黑點處),箭頭代表實體間的「活動」(activity)。機制運作起來,有起點、有終點。上層橢圓邊界表示現象範圍,下層橢圓邊界表示機制運作範圍,因為機制是產生現象的原因,所以上下兩層橢圓的邊界會一致(見虛線處)。
圖1:機制示意圖,上層是現象,下層是機制。機制由實體(黑點)和活動(箭頭)組成。機制的範圍和現象的範圍互相吻合。(圖重製於 Craver and Darden 2001)
格林蘭在 1996 年最先提出的機制概念是「在行為底層的機制是一個複雜系統,由…許多局部根據直接的因果定律進行互動...」(Glennen 1996: 52) 我們可以看到,在這個機制概念版本裡,仍依賴「定律」作為機制規律行為的因果來源。然而,格林蘭後來受到米契爾 (Sandra Mitchell, 1977) 與伍德沃 (James Woodward, 2000) 的影響,同意用「通則」(generalizations) 比用「定律」更適合生物現象,所以將機制概念裡的「因果定律」(causal laws) 替換為「與變化有關的通則」(change-relating generalizations),用通則說明機制局部間所產生的性質變化,也就是後來在2002年提出來的機制概念內容(如上 (I))。第二個機制概念是麥契曼、達登、與克瑞弗的「二元論」(dualism),他們以「實體」(entity) 和「活動」(activity) 作為機制最主要的內涵,如 (II) 所示,機制是由實體與活動所構成的組織,而組織運作起來會產生規律的變化。第三個機制概念是貝克托和亞柏漢森提出來的,內容看似延用了 MDC 的二元論,只是把「實體」換成「局部」、「活動」換成「運作」(operating),但貝克托與亞伯漢森後來的作品以及後來和其他新機制論者合作的文章裡,都更關注機制運作的動態性,而不再把機制概念停留在靜態或線性式 (linearity) 的理解裡 (Bechtel and Abrahamsen 2010; Kaplan and Bechtel 2011; Kaplan and Craver 2011; Levy and Bechtel 2013)。[3]
當我們對機制概念有初步理解後,接下來要認識機制說明的被說明項,也就是現象,因為現象是科學實作最先被感官觀察到的對象,生物學家欲發現機制,必須先對現象要有所了解,才能發現底層的機制內容,形成機制說明,最後建立說明項。
生物學家發現機制時,大都會先從可觀察的現象開始著手,她必須先確定現象表現的範圍,從有限的範圍內進行研究。當研究者對現象了解地越多,就越有利於機制發現。克瑞弗和達登對現象提出至少三種條件:第一是「觸發條件」(precipitating conditions),就是使現象從靜止不動或未顯現的狀態,變成開始動作或顯現 (manifest) 的條件,也稱「起始條件」(set-up condition)。第二是 「抑制條件」(inhibiting conditions),就是使原本正在運作的現象,變停止或趨緩的條件。第三是現象的「調整條件」(modulating conditions),是使正處於某種運作狀態的現象(正在增強或減弱趨勢),往研究者想要改變的方向予以調整(使變趨緩或便增強)。如果科學家能掌握這三種條件,就能透過干預或操控現象,探查機制內容 (Craver and Darden 2013: 56-58)。
當生物學家了解關於現象各種條件後,會用「拆解」或「解組」(decomposition) 的方式來發現機制內容。首先區分出較大的機制區塊將其「定位」(localization),然後再針對區塊內的小區塊進行解組與定位,如此程序重複下去,直到把機制內所有成分的相對位置、彼此間的連結性 (connectivity)、以及各自有什麼功能都弄清楚了,機制發現的工作也就完成了。貝克托和理查森認為,這和我們平常在理解真實機器的內部構造的認知過程很相像,都是透過化繁為簡的方式來理解事物的複雜性 (Bechtel and Richardson 1993: 17)。
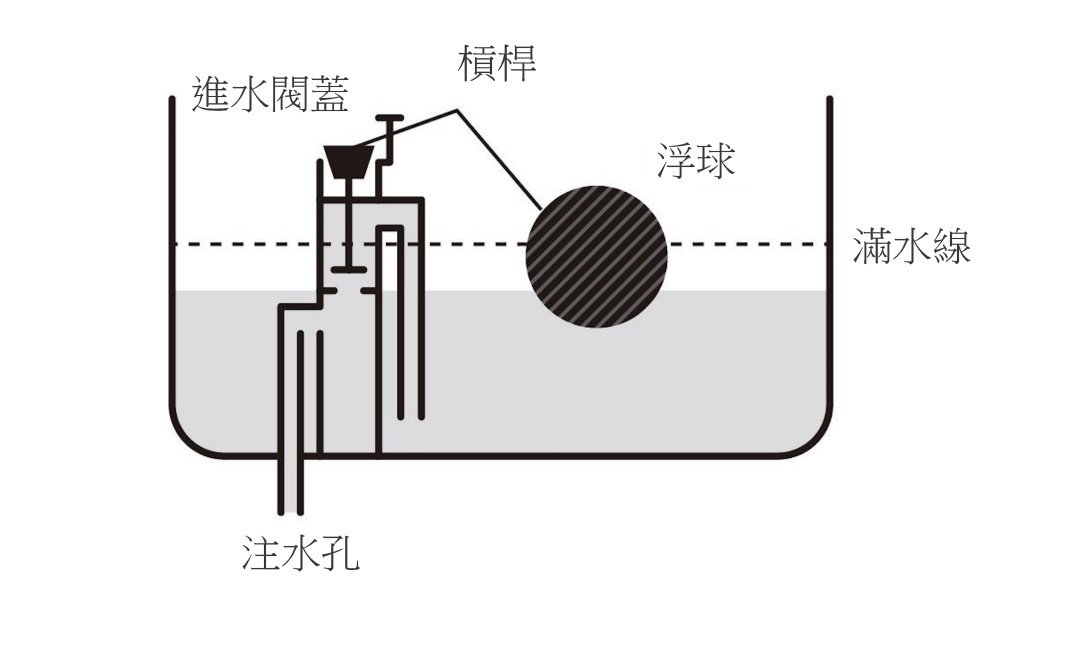
不過,格林蘭認為貝克托與理查森對機制成分間的連結性沒有交代清楚,於是他建議另一種強調局部如何「互動」(interaction) 的描述,也就是「各局部是根據因果律互動而產生機制行為」(Glennan 1996: 52) 格林蘭以抽水系統為例,抽水系統裡的所有局部包括水、浮球、控制水流進入或停止的閥蓋以及連結浮球與閥蓋的槓桿。當我按下啟動按鈕,系統的水就會開始流出水缸,水位會開始下降,浮球也跟著下降。當水位下降到某個低水位時,與浮球連結的槓桿會因為物理位能的變化使閥蓋打開,此時系統的水會開始注入水缸,此時水位開始上升,浮球也跟著漂起。當水位上升到某個高水位時,浮球槓桿會使閥蓋關閉,此時水停止注入水缸。如果我又按下按鈕,這個抽水機制就會反覆運作(圖 2)。抽水系統的運作完全符合格林蘭對機制成分間互動的描述。
圖2:格林蘭的抽水系統
2002 年,格林蘭把「互動」調整為「一個局部的性質變化引起另一個局部的性質變化」,同時也把「因果定律」改為「直接、不變、與變化有關的通則」(Glennan 2002: S344)。回到抽水系統的例子,「與變化有關的通則」就是浮力原理,「一個局部的性質變化引起另一局部的性質變化」指的是「當水位下降到低水位時,槓桿會使閥蓋打開;當水位上升到高水位時,槓桿會使閥蓋關閉」,「水位從高變低」的性質變化引起「閥蓋從打開到關閉」的性質變化。
接著是麥契默、達登、和克瑞弗所提出的二元機制組織。麥契默三人主張,機制是由「實體」和「活動」所組成的二元體 (Machamer, Darden, and Craver 2000),實體是參與活動的東西,因為具備某些性質,所以能產生活動;活動是實體所做的事情,把實體和活動組織起來,就能使機制運作發揮特定功能。讓我們借用一段取自生化學教科書的機制描述來看 MDC 式的機制組織:
DNA 複製機制從一組雙股螺旋開始,結束時會產生兩組雙股螺旋。啟動蛋白負責啟動複製機制,它和 DNA 複製起始點結合後,解螺旋酶就會扭開雙股螺旋,形成兩條單股螺旋,也就是複製叉 (replication fork)。分離後的單股螺旋就充當合成模板,上面有許多核苷酸 ,DNA 聚合酶與之結合後,沿著複製叉的移動方向前進,按照互補鹼基配對規則,一一配對核苷酸,產生一條新的雙股螺旋 DNA。[4]
這是一段描述 DNA 如何複製的機制內容,也是很典型的二元論式機制描述,整個機制有明確的起點和終點,實體包括啟動蛋白、DNA 複製起始點、解螺旋酶、DNA聚合酶,活動有結合和扭開。每組的實體和活動會引發下一組的實體和活動,實體和活動所產生的「生產性變化」(productive change) 會使機制保持「生產連續性」(productive continuity) (Machamer, Darden, and Craver 2000: 3)。
MDC 認為,活動至少有四種類型:「幾何機械的」(geometrico-mechanical)、「化學鏈結的」(chemical-bonding)、「能量的」(energetic)、以及「電磁學的」(electromagentic),其中幾何機械式的活動,有打開、關閉、嵌合、鎖上等活動;化學鍵結有吸引、排斥、擴散、斷裂等活動;能量方面活動,像是滲透、擴散(通常以熱力學作為動力來源)等等;電磁學方面的活動是指在電磁學層次所產生的電磁效應,通常這類的活動會被認為是機制結構中最底層的活動,因此 MDC 稱為「觸底 (bottom-out)」。雖然 MDC 對活動做出這四種類別,但她們也聲明,這些區分只是為了幫助理解活動可能含有不同層次,實際上,這些層次之間倒沒有絕對明確的界線存在,比如有些實體在產生幾何機械的活動時,可能會同時產生其他較低層次的活動,如影響到化學鍵結反應,所以,這些區分是為了認知上的目的。
最後,實體和活動的組織方式可從兩個面向來看,分別是空間上 (spatial) 和時間上 (temporal) 面向。空間上的組織包括局部或成分的形狀、位置、空間劃分 (compartmentalization)、局部之間的邊界 (boundaries) 等等,而時間上的組織是指活動的順序、速度、頻率、以及歷時多久 (durations) 等性質。如果我們把機制裡所有實體和活動,還有所有相關的空間與時間組織都組織起來,就有可能會組成很大範圍或層次很多的機制組織。我們可想而知,越複雜的機制組織運作,便能形成越複雜的生物現象,例子可見第三節:機制整合。
傳統邏輯經驗論以涵蓋律模型 (covering-law model) 作為科學說明的主要模型。涵蓋律模型是由普遍定律 (general law)、先行條件 (antecedent conditions)、以及待說明現象的描述語句所組成。普遍定律是說明項 (explanans),待說明現象描述句是被說明項 (explanandum)。邏輯經驗論者認為,科學說明就是從普遍定律、搭配先行條件,邏輯演繹出待說明描述句,就是「科學說明」(scientific explanation) (Hempel and Oppenheim 1948;Hempel 1965)。然而,一般生物學並不像物理學是由定律構成,那要如何套用涵蓋律模型來進行生物學說明呢?僅管古典遺傳學有孟德爾定律,例如第一條定律是分離律,以「定律」為名,但生物學家也發現到有些基因的傳遞行為似乎不會「遵照」定律的內容,那麼要說孟德爾定律不是定律嗎?這似乎也不對,因為絕大部分的基因行為確實符合了孟德爾定律的描述,那怎麼辦呢?新機制論為解決這個問題,提供一個全新的說明模式來說明生物現象,那就是建立「機制說明」(mechanistic explanation)。機制說明在絕大部分的情況下具有規律性與普遍性,但不像物理化學定律那般要求必須毫無例外。機制說明容許有例外發生,因為生物現象有時很奧妙,會有突變、發生變異的時候,而機制說明正好符合這類情況。
再者,傳統涵蓋律模式提供從前提到結論之間的演繹過程與結果,但沒有說明現象發生的原因 (cause),例如細胞為什麼會新陳代謝?神經元為什麼會去極化?涵蓋律模式能告訴我們的是依據某條定律,加上某些先行條件,能演繹出某現象語句,但這真的有說明到現象產生的原因嗎?新機制論認為沒有,真正造成現象的原因是隱藏在現象底層的機制,因此,若我們想知道現象的起因,應該不是去尋找定律與現象語句之間的邏輯關係,而是要去尋找隱藏在現象底層的機制 (Glennan 1996: 64; Machamer, Darden, and Craver 2000: 6; Machamer 2004: 28; Darden 2008: 964)。
此外,機制說明還有一個特色,就是強調「特異性」或「專一性」(specificity)。相較於邏輯經驗論重視定律是否有涵蓋到所有個別現象的普遍性 (generality),包含要能預測未來還沒發生的事件也要如定律描述那般,新機制論則認為,由於生物現象很容易出現變異 (variant),實際層面上無法被普遍化,所以,生物學家在建立機制說明時,不會把普遍性列為首要目標,新機制論者容許機制說明可以因生物對象之本質的不同而有特異性 (Bechtel and Abrahamsen 2005: 421-422)。或許有人會問:不具有普遍性的說明還是科學說明嗎?對於此問題,新機制論者會這樣回答:生物學家在最開始發展機制說明時,會先提出一個原型 (prototype) 或模型系統 (model system),然後依據研究對象的本質,發展出符合它的機制說明 (Bechtel and Abrahamsen 2005)。例如,在 1950 年代以前,神經科學家最常選用的研究對象是烏賊,因為烏賊的神經軸突 (axon) 比較粗,容易研究,但到了 1970 年代,科學家則開始改用電鱙魚,用來研究乙醯膽鹼 (acetycholine) 和神經接受器如何傳遞訊號等現象。這表示,為研究生物現象發現機制,科學家會先從一個原始或素樸的研究原型或模型出發,逐步發展出最適合的機制說明(洪純道與孫興祥,2000)。在分子生物學也類似的例子,如蛋白質合成機制也會隨生物物種的不同而有所不同,如原核生物和真核生物;或相反,原本兩個差別很大的物種,它們的記憶機制卻十分相似。因此,新機制論者建議我們用「家族相似性」(family similarity) 的概念來理解各種機制說明間的相似與相異性 (Bechtel and Abrahamsen 2005: 438; Glennan 2017, ch. 3)。
機制說明還有一個視覺上的説明優勢,就是「圖像推理」(diagrammatic reasoning)。生物學家常在機制說明裡放一些由圖案和箭頭構成俗稱的「動畫圖」(animation),圖裡會簡單標明實體名稱以及活動內容,箭頭會表示活動作用的對象或活動的時間順序,這些由非文字性的表徵內容將形成一種「視覺空間式的表徵」(visuospatial representation),帶領認知者隨著圖與箭頭啟動某種「心靈模擬或動畫」(mental simulation or animation),幫助認知者理解或想像機制整體動態運作的情形,特別是近年來,生物學研究愈加依賴電腦模擬使資料圖像動畫化,所以,圖像推理在生物發現機制研究中,扮演的角色愈來愈重要,從在機制説明裡提供視覺推理優勢,轉而作為研究者發現機制的重要知識論資源 (epistemic resource) (Bechtel and Abrahamsen 2005; Glennan 2017, ch. 3)。
上段提到,科學家在開始發展機制說明時,會先建立一個較基礎性的原型或模型系統,其實就是可以發展為完整機制描述的初始模型,也稱「機制模型」(mechanistic model)。先從一個抽象的模型出發,慢慢增添各式細節,使抽象模型成為完整的說明項,這類理論發展的模式跟早期科學哲學討論理論與模型的關係類似,但就像前面提過的,傳統科學理論是以定律為核心,但生物學沒有定律,生物學家傾向用機制來做說明項,那生物機制與模型的關係是什麼?為釐清這個問題,我們有必要簡單談一下機制與模型的關係。
談到「模型」,科學哲學從 1980 年代開始就有很豐富的討論,本文從薩普 (Patrick Suppes, 1977) 與凡弗拉森 (Bas C. Van Fraassen, 1980) 所形成的「語意觀點」(semantic view) 開始談起。簡單地說,科學理論的核心就是構成理論的語意結構 (semantic structure),語意結構是表徵理論的邏輯模型。一個科學家若能為現象建立一組抽象的邏輯結構,那基本上,他就提供了一個新理論的模型。因此,在語義觀點下,理論即模型 (theory as model),而模型代表了理論。不過,我們不打算在此進入模型與表徵對象,也就是抽象模型與具體世界之間如何判斷是否相似、以及模型真的有確實表徵到世界等複雜問題[5],在此我們僅聚焦後來由美國科學哲學家吉爾瑞 (Ronal N. Giere) 所發展出來的認知模型進路,來銜接討論機制說明與機制模型的關係。我們將問:新機制論如何定位模型在機制裡扮演什麼角色?以及在實作上,生物學家如何藉由機制模型,去發現、說明、甚至是預測某個生物系統?我們先談一下吉爾瑞的認知模型理論。
吉爾瑞 (Giere 1988, 1999, 2004, 2006) 認為,「模型」是科學家用來表徵真實世界的抽象結構,但模型不一定要是像數學方程式類型的定律系統,換言之,吉爾瑞的模型不是邏輯命題式的表徵,而是科學家在心智上,對真實世界進行「抽取」與「理想化」的思考工具,模型是科學家之於真實世界的一種「心智表徵」(mental representation)(陳瑞麟,2010,頁329)。我們可以以地圖來理解之,一張地圖就好像是一個模型,以抽象的圖形表徵了真實世界的某個面向,以台北捷運路網圖來說,圖形內容表徵了台北捷運系統的某一面向,如黃線各站的相對順序,古亭站在東門站與頂溪站中間,但這只表徵了台北捷運的其中一個面向(黃線、古亭站的前後站),假若此刻某位地圖的使用者想知道的是,從古亭站到東門站實際上間隔多長距離,那就需要另一份地圖(模型)來表徵這方面的結構關係。因此,模型是認知者認識世界的一種媒介。對科學家來說,藉由認知模型,可為真實世界系統建立相對應的科學理論,如物理學家透過力學模型,建立古典力學。那以生物學來說,生物學家就是透過機制模型,建立機制說明。
從新機制論的觀點來說,相關於機制的模型主要有兩種,一種是表徵機制內部的「機制模型」(mechanistic models),另一種是表徵機制產物的「現象模型」(phenomenal models)。前者描述了機制行為是如何由成分及其活動組織產生出來的,而後者描述了機制運作後的產物,也就是機制行為 (Glannan 2005)。對克瑞弗與克普蘭來說,這裡的區分是重要的,因為兩者之中,只有前者具有因果說明力,後者沒有。他們以神經生理學為例,論證機制模型是真正承載機制說明的載具 (vehicles),提供了機制組織由什麼實體,產生什麼活動,所以造成什麼現象,這樣才算是對生物現象提供機制說明 (Craver 2006; Kaplan and Bechtel 2011; Craver 2011; Kaplan 2011)。
機制模型如何進行機制說明呢?剛剛提到,模型隨著使用者的需求不同,會在選取表徵的面向與程度上有所不同,特別是對達登與克瑞弗來說,機制發現必須從抽象的「機制構想」(mechanism sketch)、到稍微有些內容的「機制框架」(mechanism schema)、再到擁有更多內容的「機制格架」(mechanism schemata),最後到完整的「機制描述」(mechanism description),這是一段從抽象程度最高,到具體程度最高的發現歷程。機制構想是科學家對研究現象的起因最初有的想法,可能是一個推測 (conjecture)、也可能是一個假說,機制框架是科學家大概設想了現象可能是由某某存有物進行某種活動所造成的推想,至於機制格架則是,雖然對機制組織已有某種程度的發現,但還是有一些未發現的部分存在,一直要到機制內所有的「黑盒子」(black box) 都被打開,填滿所有的未知細節,才算完成完整的機制說制,這一整套過程,達登說是「機制例示化」(mechanism instantiation) 的過程(本文第四節會有更詳細的說明)。
那模型到底在機制說明扮演什麼角色?對達登來說,上述每個機制階段都是機制本身的一種模型,發現機制就是從抽象、未知成分很多的「可能如何」機制模型 (how-possibly models),逐步發展為整體已知、具有完整描述的「實際如何」(how-actually models) 機制說明 (Machamer, Darden, and Craver 2000; Darden and Craver 2002; Craver 2007; Darden 2007: 145; Craver and Darden 2013)。但是,格林蘭認為我們不需要對機制模型進行這些區分,因為在實作上,不同機制模型間的界線不知道要畫在哪裡,他以語言認知的案例來論證,科學家所使用的機制模型從頭到尾其實只有一個,只是這個模型在科學家發展和檢驗的歷程中,會被不斷地修正與精練,直到成為適當的機制說明 (Glennan 2005; 2017 ch.3)。[6]
前面提到,新機制論者先驅溫薩特說,生物學家的工作不在於研究生物學是否可以被化約至物理與化學,而在於發現機制 (Wimsatt 1972: 67, 1976: 671, 1980),這裡說的化約是「理論化約」(theory reduction)。理論化約是科學哲學中處理理論關係的重要議題,接下來我們就要來談,新機制哲學如何重新看待生物學裡的理論關係?和傳統科學哲學理論化約論的觀點有何不同?最大的不同應該是在於傳統的理論化約論很容易對科學得出一種「科學統一」(the unity of science) 的圖像,而新機制論則是從系統性地縱覽生物學機制乃來自各領域,並且處於互為局部與整體的關係,因此,新機制論為生物學理論關係發展出各領域生物機制是自主卻不斷裂、可連結但多元的整合 (integration) 關係,不是傳統從高層理論往低層理論化約的線性關係。可是,這樣非化約、非線性的觀點很快就會招來新的質疑,例如:整合跟統一 (unification) 一樣嗎?既然是整合,又為何會得出生物理論為多元論 (pluarlism) 的結果?讓我們一一解答。
談化約,我們就要再回到邏輯經驗論。邏輯經驗論主張用涵蓋律模型說明現象,他們也用一樣的模型來說明科學進展,將舊理論視為模型中的待說明項,新理論是說明項,如果新理論的方法論可以邏輯演繹出舊理論的方法論,表示新理論「化約」(reduce) 舊理論,或反過來說,舊理論被新理論化約了。因為可以化約,表示新理論的方法論比舊理論的方法論更簡潔、或說明力更強、推廣性較高等優點,基於此,新理論得以取代舊理論,推動科學往前進展。以物理學來說,牛頓的重力定律化約了伽利略的自由落體論與克普勒的行星三大定律,統計力學化約了熱力學,以此達成物理學的進步。在這樣化約觀點下,理論上所有的科學應該都可以被化約至物理學,生物學化約到化學,化學再化約到物理學,如此一來,整個科學整體將呈現以物理學為基礎的化約、統一圖像——所有的自然現象最終都可以透過涵蓋律模式,由物理定律說明之 (Nagel 1961)。
然而,這一套模式真的可以用來處理生物學的理論關係嗎?我們可以用相同模式來看待生物學進展嗎?事實上,從生物學邁向分子化之後,生物哲學家也開始思考這個問題:古典遺傳學和分子生物學的關係也是上述的理論化約關係嗎?古典遺傳學真的可以化約為分子生物學嗎?古典遺傳學描述的對象明明是細胞裡染色體的遺傳活動(染色體複製和分離),但分子生物學描述的是分子層次的 DNA 活動(基因複製、轉錄或轉譯等),層次不同的生物學內容,可以套用涵蓋律模式、建立邏輯演繹關係嗎?我們把主張可以建立化約關係的觀點稱為「化約論」(reductionism)。化約論認為,分子生物學在方法論上確實比古典遺傳學的方法論更為簡約與清楚,而且較有訊息內容,讓我們能以較微觀的尺度理解生物世界,所以,分子生物學可以作為「化約理論」(the reducing theory) 來說明古典遺傳學(被化約的理論)。雖然分子生物學不是由定律所構成,無法建立嚴格規定下的邏輯演繹關係,但化約論者建議我們改用「可連結性」(connectability) 取代「可演繹性」(derivability),也就是在古典遺傳學的語詞(如古典遺傳學的「基因」)和分子生物學的語詞(如分子層次的「基因」,即「DNA 片段」)建立一種類似橋律的連結,使兩組理論的語詞能以系統性的方式連結起來,這樣兩個理論的關係就是理論化約關係 (Schaffner 1993)。
反對化約論的立場稱為「反化約論」(anti-reductionism),反化約論者針對上述化約論的可連結主張提出「不可連結」(unconnectability) 的反駁,他們的想法是,古典遺傳學不論在基因語詞的內涵或基因語詞指稱的對象都與分子生物學的基因語詞內涵和指涉對象的差異都非常大,以致無法在兩個理論之間建立系統性的連結。「不可連結」可從三個方面來看:第一,胡爾 (David L. Hull) 和羅森伯格 (Alexander Rosenberg) 指出基因和基因的表現是多對多的關係,所以無法建立系統化的關係。由古典遺傳學所指稱的某個性狀(如果蠅的紅眼),實際上可能是由多個分子機制產生,而且影響的「路徑包含許多其他、含混、與互相依賴的路徑。」(Rosenberg 1985: 101)。因此,古典遺傳學的語詞和分子生物學的語詞是複雜的「一對多」或「多對多」的因果關係,所以無法建立系統化和可理解的對應關係 (Hull 1974: 39)。第二,基因概念的內涵差距太大,我們無法用分子生物學的「基因」概念去設想古典遺傳學的「基因」概念,反之亦然。古典遺傳學對基因的設想是粒子式的基因概念,每個基因獨立負責每個性狀,而且,古典遺傳學的基因跟它在染色體上的位置關連非常密切,反觀分子生物學所使用的「基因」概念僅僅是「一段為產生某種特定序列之多胜肽的編碼結構」,如此可見,兩理論的「基因」概念差距頗大,無法建立可操作的對應連結。第三,基雀爾 (Philip Kitcher) 從理論自主性的觀點來看,認為我們應該讓古典遺傳學和分子生物學各自保有它們屬於該層次的自主性,維持兩學科的獨立性。換言之,應該讓染色體配對和減數分裂等現象保留給古典遺傳學或細胞學負責說明,而分子生物學就專門處理分子層次的生物現象就好。我們不需要把所有的生物說明都換為分子式的說明,或許讓專屬層次的理論來說明該層次的現象會更加適合 (Kitcher 1984)。以上,就是反化約論主張古典遺傳學與分子生物學的關係是非化約關係的理由。
關於古典遺傳學和分子生物學的關係,達登主張,無論是古典遺傳學或分子遺傳學,都是在調查不同、但是連續、在同一系列的遺傳機制,意思是說,雖然古典遺傳學的研究對象是染色體獨立分配和分離的機制,而分子生物學是研究 DNA 複製的機制,表面看去,兩者分屬不同層次,反化約論認為無法化約。然而,達登認為兩者機制雖然層次不同,但可以連結。當分子生物學家發現蛋白質合成機制和基因表現路徑時,其實是在為古典遺傳學的染色體複製機制,提供更微觀與詳細的機制內容(圖三)。換言之,分子生物學是以提供較細部的機制內容的方式與古典遺傳學連結。分子生物學所發現到的內容,會解開古典遺傳學尚未解開的謎題。這樣的連結,對古典遺傳學來說,分子生物學的發現填補了其理論未解的部分;對分子生物學來說,古典遺傳學的發現提供了在它以外的更大知識脈絡。無論古典遺傳學發現的機制,還是分子生物學發現的機制,都是遺傳整體的一部分。因此,兩門生物學應是「局部與整體」的連結關係,不是誰被誰化約的化約關係 (Darden 2005: 351)。
圖3:古典遺傳學和分子生物學機制是連續且整合的遺傳機制,二者呈現部分和整體的關係。圖中白色橢圓代表古典遺傳學機制細胞內染色體的各種活動,包括交換、獨立分配、減數分裂等。黑色方框代表分子生物學所發現的基因複製和蛋白質合成機制。白色橢圓和黑色框框都可以被整合在一個大的機制系列裡,不是由上而下的化約關係。(圖重製自 Darden 2005)
對新機制論來說,一個生物學領域為另一個領域提供機制內容代表兩領域是互相連結的關係,但連結不會影響領域的獨立性。對達登來說,所有的生物學領域都可以各自有獨立,又可以互相提供機制資源。如此一來,我們就可以不用依賴化約論來理解生物學的理論關係以及進展。不同領域的生物學家一方面調查專屬層次的機制內容,另一方面可以提供其他層次所需要的機制說明。各領域的機制內容可以被整合成一個在時間上連續、不斷裂的遺傳機制,形成一種「機制整合」(mechanism integration) (Darden 2005: 349)。除了達登,還有克瑞弗也是用機制整合觀點重新處理神經生理學的機制層次。克瑞弗主張,透過對不同層次的機制進行整合,可以為神經科學提供一個馬賽克式的整合圖像。他與達登稍有不同的是,克瑞弗處理的是機制「階層」間的整合,達登處理的是領域間的整合。
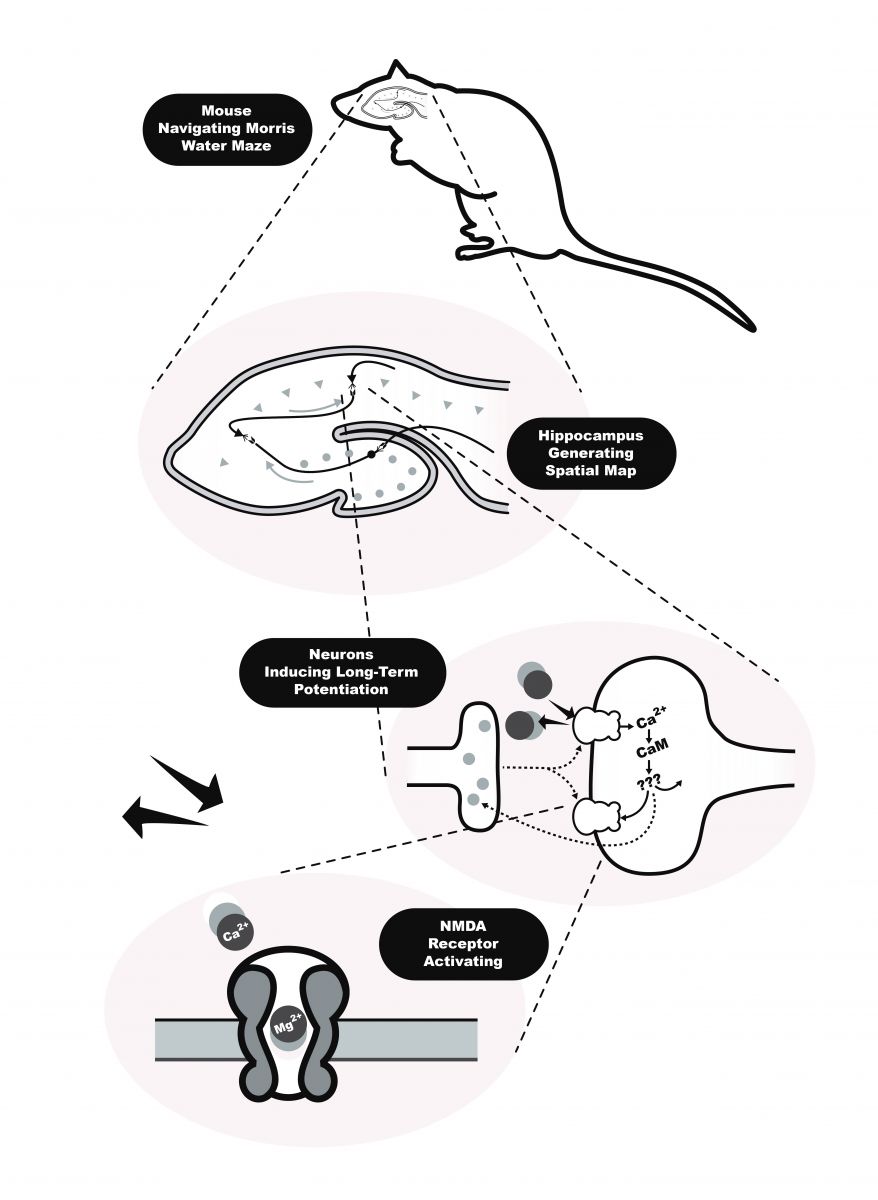
克瑞弗以動物學習記憶為例,學習記憶涉及至少四個層次的機制:「生物—行為」、「海馬迴—訊號傳遞」、「神經元—活動」、「分子—動力」。這四個層次表徵了不同尺度的機制活動,第一層「生物—行為」層次,研究動物如何儲存、增強或降低學習與記憶能力;第二層「海馬迴—訊號傳遞」層次,研究動物大腦裡海馬迴和內側額葉裡神經活動的變化;第三層「神經元—活動」層次,研究神經細胞、傳導物質、和各種離子間的活動(包括釋放、結合、擴散、通過與行動電位變化等);第四層「分子—動力」層次,研究更底層的實體,如 NMDA、AMPA 接收器、麩胺酸、鈣離子與鎂離子是如何活動(活化、吸引、阻擋、磷酸化、水解等),但這已深入到生化學、藥理學和電生理學的層次了。一般而言,較高層次的機制常常是科學研究的起點,它提供研究者機制框架、指引研究方向。對克瑞弗來說,每層次的機制內容就像一片片的馬賽克圖片,科學家透過發現各層次的機制內容拼貼出一幅跨層次 (interlevel) 的整合機制,最後形成巢狀網路式 (nested networks) 的大機制。
圖4:動物學習記憶的四個層次(圖重置自 Craver 2002)
從上面兩個例子中,我們不難看出,無論是生物學還是神經生理學都是「跨領域」或「跨層次」的整合式說明。整合式說明表徵機制間存在部分與整體的關係,低層次的機制是高層次機制的一部分。各層次的機制都有屬於該層次的機制內容,認知者可依認識興趣在不同層次裡尋找欲得知的機制內容。整體而言,機制說明就像每片馬賽克圖片一樣,各自獨立存在,無法被其他圖片取代 (Craver 2001, 2007, 2012)。為了發現跨領域或跨層次機制,科學家有時必須先往其他領域找資源,有時往較低層次,有時往較高層次,然後將不同層次的內容整合起來,才能看到機制全貌。換言之,生物學的理論關係未必只有走向化約一條路。在新機制觀點下的理論關係,呈現一幅包含局部 (local) 且多元 (pluralistic) 的圖像 (Darden and Cain 1989)。[7]
我們前面談了機制概念、機制說明、以及用機制整合觀點來看生物學理論間的關係。現在我們要看機制是如何被發現的?這是科學發現 (scientific discovery) 的老問題,邏輯經驗論曾提出「發現脈絡」(the context of discovery) 與「證成脈絡」(the context of justification) 的區分。但他們主要討論的都是證成脈絡,發現脈絡反而談得不多。原因可能是,當時的邏輯經驗論者認為,科學家在提出一個新假設或新理論的過程中,推理的過程應該是相當隱微、迂迴、或變動不定的,如果是這樣的話,那哲學家很難為此發展出一個相對穩定的發現模式,所以就把「發現」的這個部分交給歷史學、心理學或社會學來處理,哲學家則聚焦分析科學假設和觀察語句之間的邏輯推理關係就好 (Popper 1959)。隨著分子生物學的發展,生物哲學家(特別是新機制論者)開始關心:那生物學的發現是如何?生物學家是怎麼發現機制的?是否共享某種固定的模式?(Schaffner 1974; Machamer, Darden, and Craver 2000; Craver and Darden 2001, 2013; Darden 1991, 2006; Darden and Craver 2002; Bechtel 2006)
對達登來說,這問題的答案是肯定的。她認為,生物學家在發現機制的過程中,有一套「發現推理」(reasoning in discoveries) 模式,她稱為「策略」(strategies)。達登說,用「策略」這個詞是受到拉卡托斯 (Imer Lakatos)「科學研究方案(或綱領)」(scientific research programme) 之「啟迪術」(heuristics) 的影響 (1978)。「啟迪術」是啟發科學家在研究過程中獲得靈感的一種推理「方法」,可是雖然說是「方法」,但不是邏輯經驗論所說的邏輯證成有明確的前提和演繹規則與結果,啟迪術比較像是,科學家在面對大量龐雜的資料時,大腦運作處理的一種推理方式。這種推理方式可能不是很嚴謹,但保持一定程度的明確性和方向性,使科學家在對資料進行分類或推理時,提出新假設。[8] 達登和克瑞弗延續這樣的思路,並貼近生物學和神經科學的實作,提出一系列詳細的發現機制策略,包括修正異例策略、建構理論或機制框架的策略、評價的策略、做實驗的策略等等 (Darden 2006)。對達登而言,這些策略不完全是描述性的 (descriptive),也不是指示性 (prescriptive) 的,發現策略是對生物學家曾經使用過的機制概念和發現機制的過程進行後設哲學分析後,所發展出來的一套方法論,它是對未來發現實作提供一種諮詢性 (advisory) 的建言 (Darden 1991: 15-17, 2006: 272)。我們無法在此介紹所有的發現策略,我們挑選其中四種做介紹:框架例示、向前或向後連結、實驗干預策略、與解決異例。
發現機制就是對現象底層的機制作出完整的機制描述。每個機制描述都是從模糊、抽象的「構想」(sketch) 開始。構想的內容通常很有限,有時只知道一個輪廓或是所觀察現象前後有些不同的特徵,真正關於機制裡的內容還一無所知,達登和克瑞弗稱為還有許多「黑盒子」(black box)。等生物學家對現象有較多的了解後,生物學家會開始提出「機制框架」(mechanism schemas),機制框架表徵了機制「有可能是」(how possibly) 某個樣子,有可能存有某種實體,但不知道它產生什麼活動,或相反,知道有某種活動,但不知道是由什麼實體所造成的,又或者是,知道現象的起始條件,但不知道終止條件或中間的歷程細節等等,有些內容已知,有些未知。此時生物學家會從她的背景知識 (background knowledge) 開始尋找可應用的資源,也就是尋找可用的「機制類型」(types of mechanisms),如研究細胞傳遞現象的科學家可能會去找,過去已發現到的傳遞機制類型,而研究基因調控現象的科學家可能會去調查,過去已有哪些已知的調控機制類型。因此,在背景知識裡尋找機制類型常常是生物學家發現機制的第一步 (Craver and Darden 2013: 67-69)。
有了機制框架後,生物學家要使框架進一步成為「機制格架」(mechanism schemata)。機制格架包含「機制很有可能 (how plausibly) 是由某些實體的某些活動所組成」的內容,那些還未被發現的實體或活動內容,達登與克瑞弗稱為「灰盒子」(gray box)。相較於黑盒子,灰盒子發現內容比較多。對生物學家來說,機制格架像是一份指引研究方向的藍圖 (blueprint),其中會應用的一個發現策略是「模組組裝」(modular subassembly) (Darden 2006)。「模組」(module) 是模型裡的一部分,一個模型可由一個或多個模組構成,好像我們在組裝汽車模型時,會有各式組件或套件,例如引擎組件或輪胎組件。模型組裝者必須先把一些小組件完成後,再以正確的組織方式黏著進車體套件裡。類似地,在發現機制的過程裡,發現者必須把組件先組裝成模組,然後再以正確的方式組織為機制框架或格架(視未知成分多寡)。當所有套件、模組、與細項成分都被正確定位後,就能讓機制發揮該有的功能。舉例來說,大腸桿菌受到誘導物的影響會啟動體內基因調控「抑制物模組」(repressor module) 生成特定酶類,抑制物模組就是由「會產生抑制物的基因」和「抑制蛋白質」兩個組件所組成。所以,當生物學家在機制假說裡放進這個模組,就可以依其設計各種實驗去測試該模組,以調查模組內其他組件的功能或性質究竟為何。
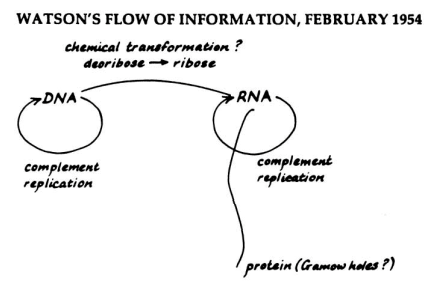
等機制格架裡所有未知的部分都被解開,表示機制已從含有黑盒子的框架,進展到含有部分未知、部分已知的灰盒子格架,最後擁有完整機制內容的「透明盒子」(glass box),那機制就被發現了,這時候科學家就可以把此機制類型應用推廣到相同類型的現象上。達登把此發現策略稱為「使框架例示化」(schema instantiation) (Machamer, Darden, and Craver 2000: 16-29; Darden 2006: 286-287; Craver and Darden 2013, ch.5)。[9]分子生物學中最知名的框架例示應該是「中心法則」(central dogma of molecular biology) 的例子。華生 (James Watson) 在 1954 年為基因表現提出中心法則,他假設「DNA 遺傳資訊會先經過某種與 RNA 有關的階段最後才變成蛋白質」(圖5)——這就是一個機制框架,對現象僅知一小部分資訊(DNA、RNA、蛋白質),其他細節都還不知道(圖中問號處)留待科學家發現。[10]為解答這些問號,生物學必須先回答中間的 RNA 是什麼實體?它在中心法則發揮什麼功能?科學家們後來發現,原來 RNA 是一種能攜帶遺傳訊息的 RNA,也就是訊息 RNA (messenger RNA),訊息 RNA 先和核糖體 (ribosome) 結合,核糖體就好像讀取傳統錄音帶的讀寫頭,會沿著訊息 RNA一面讀取遺傳資訊,同時將遺傳資訊依序排列出來組成蛋白質,這才解開機制框架裡的問號。這一方面需要生化學家詹美尼克 (Paul Zamecnik) 從能量的角度,回答細胞製造蛋白質的能量來源以及中間能量要如何轉移等問題,另一方面需要分子生物學家站在遺傳資訊的角度,回答 DNA 遺傳資訊究竟是如何轉移成為為蛋白質的問題。換言之,科學家提出框架後,就要開始解開框架裡的黑盒子,填滿機制過程裡的所有間隙 (gap),才算完成機制發現 (Darden and Craver 2002)。
圖5:中心法則示意圖。華生當時假設生物遺傳的「訊息流」(flow of information) 自 DNA 互補複製 (complement replication) 後,透過某種化學轉型,傳遞到 RNA、再到蛋白質,但是是怎樣的化學轉型當時還不知道(圖上方的問號),而且遺傳訊息是在哪裡形成蛋白質也不清楚(圖右下方的問號)。(圖取自 Darden 2006: 31)
機制是現象出現的原因,機制必須在現象開始出現到停止這中間保持「連續性」(continuity),現象才會持續顯現。連續性不僅使現象持續顯現,也讓機制從初始階段到終止階段都保持運作,直到機制停止,現象才會跟著也停止 (Craver and Darden 2001)。因此,生物學家必須把機制裡每個階段的實體和活動找出來,包括連接不同階段的實體或活動,有時生物學家會先知道活動內容但不知道實體,此時生物學家就會從活動內容去推理可能是由哪種實體所造成的,因為它具有某種性質可以產生該種活動,有時生物學家是先知道實體但不知道活動,此時生物學家就會從階段前後去推理可能是什麼活動內容,才會讓前後階段件連結起來,這就是新機制論者說的「向前或向後連結」的策略。
回到中心法則的例子,不同領域生物學家解開框架裡不同的黑盒子或為間隙提供不同的資訊,其實就是在連結機制生產連續性。生化學家發現介於胺基酸 (amino acids) 和多胜肽鏈之間的能量中繼物 (energy intermediates)(即發現實體),分子生物學家發現遺傳資訊如何從 DNA 傳遞到 RNA 再傳遞到蛋白質(即發現活動)。前者發現機制從起始點到終止點的能量流 (energy flow),後者發現機制的資訊流 (information flow)。「分子生物學家從 DNA 往前推理,同時生化學家從完成的蛋白質往回推理;他們帶著發現到的不同種類 RNA 及其角色在機制中間點會合。」(Darden 2006: 288) 兩領域的科學家都必須在機制框架裡,反覆地向前與向後尋找實體和活動,唯有把所有階段的實體、活動、組織都整合為完整的機制描述,機制才算完成發現。
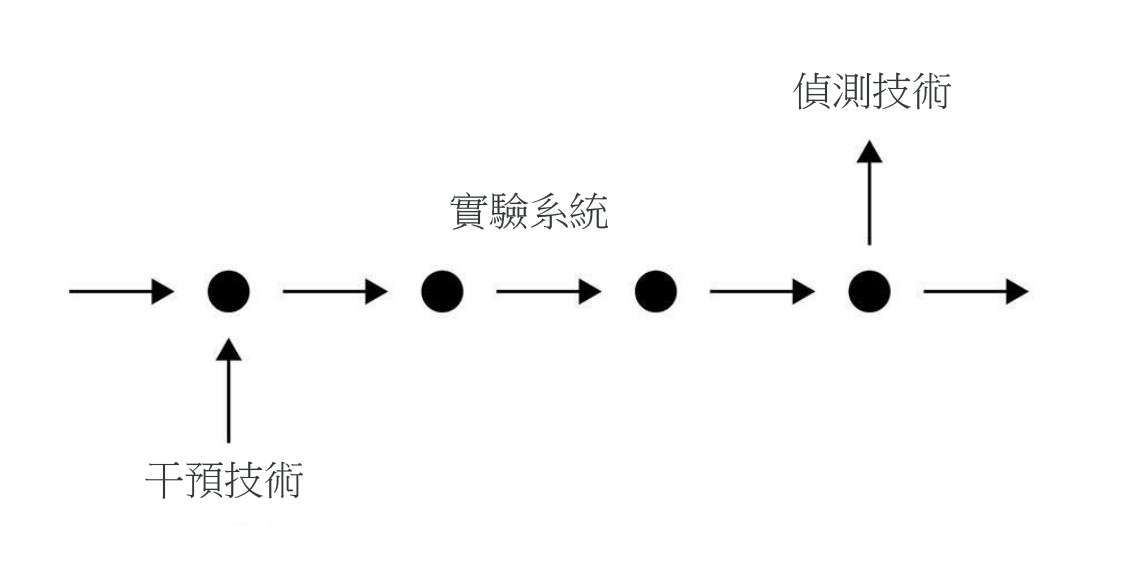
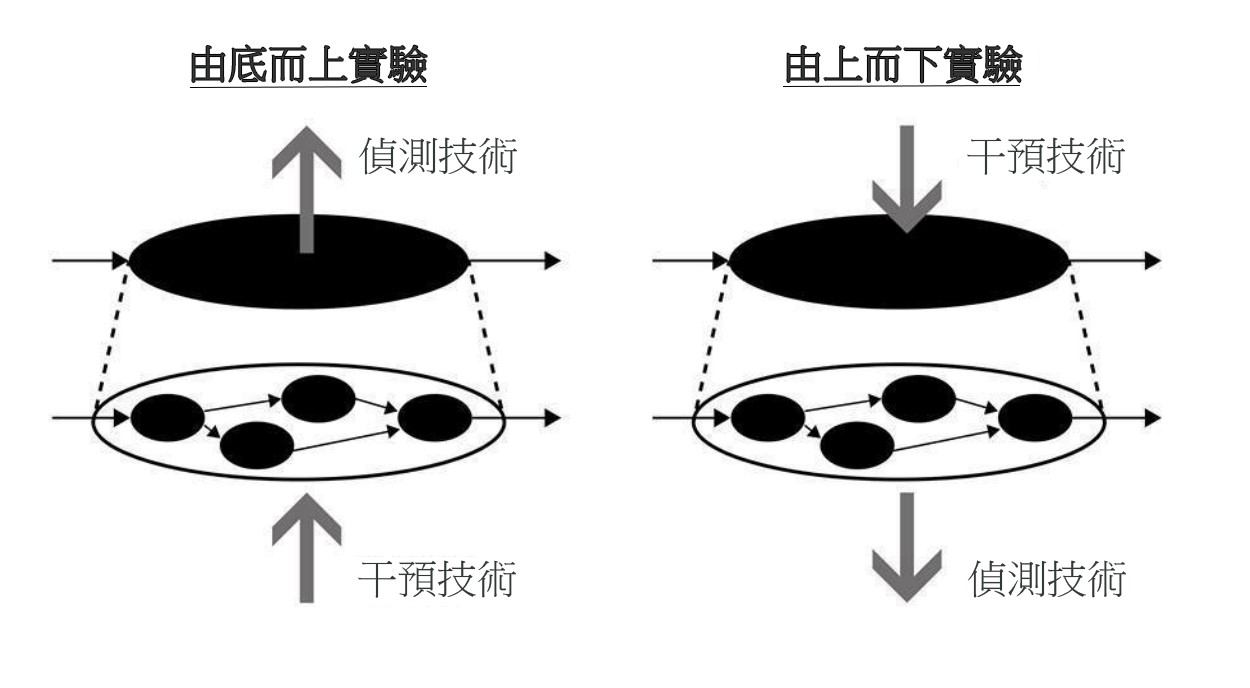
在實驗方面,新機制哲學延續培根 (Francis Bacon) 實驗哲學 (experimental philosophy) 的精神,將實驗視為發現新機制的重要工具。如前所述,生物學家會先對現象起因提出一個機制框架,框架內含有許多未知的成分。生物學家為揭開未知成分,會設計各種實驗測試框架所假設的內容是否正確,然後從實驗結果去修改或調整機制框架,直到框架變成完整的機制描述為止 (Machamer, Darden, and Craver 2000: 17)。為理解實驗行為裡的「干預」(intervention),克瑞弗和達登引用英國哲學家伍得沃德 (James Woodward) 的干預理論 (interventionist theory),透過外在操縱因果變項,使原因和結果兩變項數值產生變化 (making a difference),來確定受干預的變項是否為結果變項的原因。克瑞弗和達登把此干預概念放入發現機制的脈絡裡,為實驗干預提出一個基本模式:「干預與偵測」(intervene-detection)(圖6)。
圖6:「干預與偵測」的基本模式。科學家在設計的實驗系統裡,對機制框架的某個部分進行干預,再對另一個部分進行干預後偵測。透過「製造差異」的過程,發現機制內容(圖重製自 Craver and Darden 2013: 122)
生物學家依據機制框架設計實驗,對其中某部分進行干預,如刺激或抑制某實體的活動使其產生變化(使反應更增強或變緩慢),然後在另一處偵測干預後的變化,然後看兩個變化值之間是否具有「因果關聯」(causal relevance)。機制裡的因果關聯包含許多層面:機制是現象的整體原因、機制成分間的因果關聯 (componential relevance)、上游階段作為下游階段的原因 (ecological relevance)、以及跨層次間的因果關聯 (interlevel relevance) (Craver and Darden 2013: 120-129)。無論生物學家欲調查的是哪一種因果關聯,都可以用干預與偵測模式。以機制作為現象原因為例,生物學家可以從機制層次進行干預,然後在現象層次偵測干預結果,這是「由底而上」(bottom-up) 的干預實驗(圖7 左邊)。反之,如果是對現象進行干預,然後偵測底層機制有何變化,就是「由上而下」(top-down) 的干預實驗(圖7 右邊)。
圖7:兩種干預方向的實驗。左邊是對底層機制進行干預,偵測現象反應,是「由底而上」的實驗。右邊是對現象進行干預,偵測底層機制變化,是「由上而下」的實驗。(圖重製自 Craver and Darden 2013: 126)
除了干預方向,還有干預效果 (effect),例如「激活」(activation),就是使原本沒有出現或處於靜息的現象啟動或顯現 (manifest) 出來,這需要科學家先找到現象的觸發或起始條件 (precipitation/set-up condiction)。第二個干預效果是「刺激」(stimulation),刺激是使底層機制的實體活動增強。第三種是「干擾」(interference),干擾是擾亂機制原有的運作狀態,使原本具抑制性質轉而開始活化或增強,或使原本具有活化性質趨於停止或減緩,這需要科學家找到現象的調整條件 (modulating condiction)。最後一種是「抑制」(inhibition),抑制是使現象趨於停止運作,這需要知道現象的抑制條件 (inhibiting condition) (Craver and Darden 2013: 56-58; 125-129)。[11]
結合干預方向與干預效果,克瑞弗和達登區分了各種干預實驗,有些是透過已知實體尋找未知活動 (by-what-activity) 實驗,有些是由已知活動尋找實體 (by-what-entity) 實驗。當欲發現的機制組織本身很複雜,調查該機制的干預實驗也就會變很複雜,變成是由多個實驗組成、含有多重干預 (multiple interventions) 的系列實驗 (Craver and Darden 2013: 133-137)。為說明不同類型的實驗,克瑞弗和達登都會舉例說明其中的干預內容,例如用十九世紀流行病學家哥德伯格(Joseph Goldberger)發現糙皮病的致病機制是由於營養不良而不是傳染病,就是透過干預病人飲食而發現的;美國藥理學家羅維 (Otto Loewi) 用干預實驗發現動物心跳頻率和交感神經機制之間的關係;二十世紀美國分子生物學家艾弗里 (Oswald Avery) 用注射病毒的干預手段發現生物遺傳物質原來是去氧核糖核酸 (DNA);美國生化學家艾克斯羅德 (Julius Axelrod) 用一系列多重干預實驗發現去腎上腺素回收機制;法國巴斯德實驗室雅克布 (Francois Jacob) 和莫諾 (Jacque Monod) 團隊設計預備實驗發現大腸桿菌乳糖操縱組等,都證明了干預實驗之於機制發現的重要性 (Craver and Darden 2013: 120-143)。
當科學家在實驗過程發現與框架預期不合的結果時,我們稱為「異例 (anomaly)。異例出現往往跟兩件事有關:一個是實驗本身,另一個是框架假設。新機制論者把跟實驗有關的錯誤稱為「實驗錯誤」(experimental and data analysis error),例如實驗設計錯誤、操作出現失誤、資料分析錯誤等等。解決這類錯誤的方法就是去調查實驗設計、操作、分析資料,找出是哪個環節出現問題。如果經過診斷,發現異例與實驗本身無關,表示是框架裡的某部分有問題 (incorrect),此時科學家就要進一步地去判斷異例的性質與種類,才能決定是部分修改 (revising) 框架內容就好,還是需要重新設計 (redesign) 一個全新的框架。
依據將對框架造成多大的修改程度,新機制論者將異例分為三大類:第一種是「怪物或特例異例」(special case or monster anomaly),第二種是「模型異例」(model anomaly),第三種是「否證性異例」(falsifying anomaly)。我們可以用光譜的概念來看這三種分類。第一種怪物或特例異例算是最不威脅原框架的異例,因為它們的出現不是因為框架哪裡出錯,而是在實驗的過程中,實驗者意外誘發不同機制出現反應,讓實驗者誤以為是出現與框架不一致的異例,但其實該意外現象與框架假設本身無關,也不是實驗出錯,而就是意外發現另一種現象的特例。舉例:有些病毒遺傳不是從 DNA 開始遺傳,而是從 RNA 開始傳遞遺傳物質,這種病毒的遺傳傳遞順序跟科學家平常所假定的遺傳順序相反(上述的中心法則),所以當實驗者一開始發現到這種病毒時,可能會先以為此病毒是與中心法則相衝突的異例,但其實不是。這些病毒只是讓生物學家認識到原來在中心法則的框架外,還有其他不一樣的傳遞方向,屬於中心法則以外的特例。
第二種「怪物異例」跟特例異例有點類似,也是一開始觀察到時,會讓生物學家懷疑是不是框架或實驗哪裡有問題,但經過重複實驗查證之後,發現原來不是框架或實驗過程有問題,而是實驗者意外發現到機制其他階段的反應。例如,古典遺傳學實驗者原本瞄準研究的是一組動物染色體在受精後(Aa 與 Aa)會如何分配?得到實驗結果是:AA : Aa : aa =0: 2: 1,這個經驗結果與孟德爾 (Gregor Mendel) 自植物得出的遺傳定律假設 1: 2: 1 不同,相較之下,有三分之一的顯性性狀 ("AA")「不見了」。這個不一致的結果是挑戰孟德爾遺傳定律的「異例」嗎?後來生物學家透過適當的實驗工具發現,原來 "AA" 配子在受精後的發育早期階段就死掉了,所以觀察者「看不到」由 "AA" 形成的顯性表現型。關於「胚胎發育早期」的現象不在原框架的研究範圍內,所以此現象不會使科學家放棄或修改原有的框架。這類的發現反而會使科學家更加確認框架為真的可能性,繼續限縮或收斂框架的假設範圍以完成框架例示化。
真正會威脅到框架的是「模型異例」。當實驗者發現疑似是異例的異常現象並對其診斷後,排除不是因為實驗錯誤、也不是特例、或來自其他階段的機制,那他們可能會很嚴肅地看待此異例,因為代表原設定的框架很有可能是錯誤的,這時候科學家不得不動手修改框架裡的內容。由於框架裡有許多模組、組件、成分,還有各種假設,包括時間性的和空間性的,因此,若要修改框架,等於是要對框架裡的模型進行修改,可能大到整組模型換掉,或只要修改某個小部分就可以,修改程度端看此模型異例對框架威脅到什麼程度。舉例:孟德爾最初設想的遺傳框架是:生物所有性狀都是獨立分配的。後來實驗發現「有一些」性狀好像會「連鎖」(linkage) 遺傳,也就是有些性狀會連帶一起傳遞到下一代。當時遺傳學家發現這個異常現象時,動搖了孟德爾遺傳框架的地位,這就是一個模型異例的例子,此異例使孟德爾不得不調整原框架裡「獨立分配」的模型。
無論是實驗錯誤還是其他類型的異例,都會讓生物學家回頭檢視框架是否正確。若僅僅只是實驗操作或分析資料出錯,就不需要調整框架。若是模型異例,就要去檢查是框架哪個部份需要修改或調整。有時生物學家心裡會有一個以上的框架,此時異例就會像「關鍵實驗」(crucial experiment),讓生物學家對不同框架進行評估和決斷,最後捨棄其中某個框架,保留另一個框架繼續研究發展下去,使框架例示為機制描述。但也有可能,異例巨大到迫使生物學家放棄整個框架,得重新構想出另一個全新類型的框架,碰到這種異例就是屬於「否證性異例」(falsifying anomaly)。無論是哪一種異例,實驗現象都會為生物學家提供發現機制的線索和資源。生物學家透過辨別與分析異例,有時調整 (fixing) 或修正 (revising) 框架,有時必須重建 (re-constructing) 另一個新的框架,直到所有的實驗現象都與機制描述真確吻合時,才算完成機制發現。
自新機制論被提出後,就受到生物學家和生物學哲學家高度矚目與大量討論。擁護新機制論基本主張的人,企圖為新機制論發展更完整的論述,包括在形上學和方法論層面。也有人嘗試將新機制論延伸應用至生物學其他領域,如演化論、發育學、系統生物學,以及社會科學領域,如經濟學。[12] 反對或懷疑新機制論的人則質疑,新機制論畢竟是從分子生物學和神經生理學等較微觀層次所延伸出來的哲學論述,如此是否適用於較宏觀的領域,如演化論,或著重跨層次研究的認知科學與社會科學領域(如電腦計算或工程科學等)。[13] 無論如何,新機制論都為科學哲學與生物學哲學翻開了新的一頁。
這股新哲學潮流在英美流行近十年後,台灣敏銳的哲學家們也已準備好與國際學者們對話。2008 年 11 月中正大學哲學系陳瑞麟教授與清華大學經濟系趙相科教授跨校合作舉辦一場跨領域國際研討會「國際生物學與經濟學哲學會議暨工作坊」(International Conference and Workshop on the Philosophy of Biology and Economy)。兩人合邀英國經濟學哲學家摩根 (Mary S. Morgan)[14]、美國生物學哲學家羅森伯格和瓦特斯 (C. Kenneth Waters) 來台與會。這次會議主題設定討論生物學和經濟學裡「模型」(model) 和「演化」(evolution) 兩個哲學概念。三年後,趙相科教授與同校哲學研究所陳思廷教授再次跨領域合作,於 2011 年 3 月清華大學舉辦「生物學與經濟學哲學國際研討會:機制與因果」(Taiwan Conference on the Philosophy of Biology and Economics: Mechanism and Causality),兩人邀請到幾位知名學者,包括新機制哲學重要人物達登、反對將機制概念用於演化論哲學家彌爾斯坦、以及其他共享此哲學興趣的國外哲學家與會。台灣方面不僅有來自生物學哲學方面的哲學家,還有來自神經科學哲學、心靈哲學、知識論、因果形上學、科學史等領域的學者們出席會議,更有關心哲學議題的物理學家和生物學家們共襄盛舉參與討論。
該會議的具體成果呈現在兩年後(也就是 2013年)出版的《生物學和經濟學的機制與因果》(Mechanism and Causality in Biology and Economics)。此本論文集共收錄「定義機制與因果」、「模型和表徵」、「重新思考生物機制和因果」、以及「跨越生物學和經濟學邊界」等四個主題,一共十二篇論文,其中台灣學者貢獻三篇。陳瑞麟的〈生物學中的實驗發現、資料模型和機制:一個來自孟德爾研究的案例〉,該文企圖爭論,有一種生物學發現至今仍未被詳細討論:實驗發現。他從古典遺傳學研究案例中發現,實驗發現來自實驗的資料模型,資料模型促使並引導生物學家發現新機制 (Chen 2013)。第二篇是台灣大學哲學系王榮麟教授的〈天擇是一種屬於族群層次的因果過程嗎?〉,此文目的在於與論文集中另一篇由美國哲學家彌爾斯坦所寫的〈天擇與因果生產性〉進行對話。彌爾斯坦之前曾爭論天擇是屬於族群層次的因果過程,但不應該用「機制」的概念來理解 (Skipper & Millstein, 2005; Millstein, 2006),王榮麟則質疑彌爾斯坦認為天擇是族群因果過程的主張,他認為若按照彌爾斯坦的說法,天擇會出現一些無法說明的情況。據此,他提供一個出自形上學考量的新觀點,使天擇的概念更為完整 (Wang 2013)。[15] 第三篇是趙相科與陳思廷合著的〈對失蹤的一億個女人提出解釋〉,這是一篇以跨領域外推 (interdisciplinary explorations) 的社會科學哲學之研究 (Chao and Chen 2013))。
2013 年克瑞弗和達登合作出版《尋找機制:生命科學跨領域的發現》一書 (In Search of Mechanisms: Discoveries Across the Life Sciences),為 2000 年以來的新機制論提供一個更完整的論述。2014 年陳瑞麟在中正大學舉辦「台日生物學哲學工作坊」(Japan-Taiwan Workshop in Philosophy and History of Biology),邀請達登與多位台灣與日本的生物學哲學學者與會。除了請達登給予一場大會演講 (keynote speech) 外,也為她與克瑞弗的新書舉辦「與作者相會」的討論會。2015 年,台灣學術期刊《科技、醫療與社會》(Taiwanese Journal for Studies of Science Technology and Medicine) 收錄達登的演說論文〈演化變遷的多層機制〉("Multilevel Mechanisms of Evolutionary Change.")(由葉筱凡翻譯為中文),以及四篇由會議發表文章改寫完成的研究論文。
另外,還有一個面向也值得一提:因果 (causation)。在新機制論興起前,英國哲學家卡特萊特 (Nancy Cartwright)[16] 就曾挑戰邏輯經驗論的「律則性因果觀」。她認為,邏輯經驗論之涵蓋律模型所依恃的前提,也就是自然律,其實是科學家為尋找大自然「原因」所設想出來的虛構物 (fiction),定律並不真實存在。她建議用「潛能」(或翻「儲能」)(capacities) 或「律則機械」(nomological mechines) 來理解自然現象真正的起因 (Cartwright 1989)。「律則機械」是卡特萊特自創的形上學語詞,用來指稱某種具有規律性的自然機械。她認為「自然機械產生自然現象」和後來新機制論認為「生物機制產生生物現象」的觀點(特別是 "MDC" 的二元論)十分類似 (Cartwright 2007: 11)。不過究竟有多相似,卡特萊特倒沒有深入地分析。陳瑞麟爬梳過去曾經指出卡特萊特的律則機械觀和MDC的二元機制觀彼此是相似的哲學論述,接著細緻地比較兩造理論之成分的異同,例如卡特萊特的「潛能」在何種意義上算是等同於 MDC 所定義的「性質」?若兩者都共享某種機械或機制是所有現象的原因,那麼在兩個概念架構裡,定律居於何種定位?經過一番對比分析後,陳瑞麟建議一個將兩種機械與機制論述予以整合的方法,使關於機制論的探討脈絡更為完整 (Chen 2017)。
如前所述,新機制哲學潮流發展至今,台灣哲學界努力地讓台灣科哲學界與國際前沿思潮接軌。受到這些研究氛圍的影響,近年也開始有延續新機制哲學研究的新生代產生。第一位是英國倫敦大學學院 (University College London, UCL) 科技史博士區曣中,她的博士論文以生物機制圖像 (diagrams) 具備何種知識論與溝通功能為研究主題。區曣中教授回台後於成功大學歷史系任教。她在研究領域方面,繼續聚焦細胞機制研究和細胞生物學之圖像證據的科哲分析。她的最新論文〈以時間與事件的關係作為建構細胞機制的表徵〉("Time-Event Relationships as Representations for Constructing Cell Mechanisms"),企圖爭論在實驗細胞生物學中,生物學家必須透過由時間和事件所組成的觀察來進行機制推理,進而為細胞現象提供機制說明。由於時間和事件在細胞生物學實作上扮演相當重要的角色,因此研究者對時間的知覺和判斷事件間的因果性也就相當重要。此文為細胞生物學之視覺化證據與機制推理提供一份新的分析 (Au 2019)。另一位是在 2017 年在中正大學取得哲學博士的葉筱凡,她的博士論文以《生命科學的機制與實驗》為研究主題,她企圖建立一個以實驗干預為中心的生命科學方法論。同年,她與陳瑞麟合作〈一個共同適用於古典遺傳學和分子生物學且以實驗為基礎的方法論〉("An Experiment-Based Methodology for Classical Genetics and Molecular Biology."),此文出版於日本科學哲學期刊 (Annals of the Japan Association for Philosophy of Science)。
新機制哲學在過去兩個世紀以來,不論是對生物學哲學或社會科學哲學領域,都帶來巨大的影響。在這則詞條裡,我們不僅看到新機制論為許多傳統科學哲學議題給出新的答案,同時也引發許多新的議題等待新機制論者繼續研究挑戰它們。無論未來新機制哲學發展如何,我們期待新機制論者繼續為生物學哲學開啟一條最貼近生物學實作的哲學思路。
[1] 關於生機論與機制論的發展可見華盛頓大學聖路易斯校區榮譽教授加倫‧艾倫(Garland E. Allen)的「機制、生物、與生機論」論文,他在文中詳細介紹生物學家從十七世紀到二十世紀間,如何理解與使用「機制」概念(Allen 2018)。此外,陳瑞麟(2009)也在「孟德爾究竟發現了什麼?一個實驗發現的典型模式」文中,陳述十九世紀裡,生物學不同理論的各條支線發展。
[2] 「定徵」(characterize)指的是為「機制」概念提供一個通用(generic)的概念內涵,不是提供充份且必要條件式的定義內容。
[3] 另外還有一位也算是新機制論的先驅—美國科學哲學家溫薩特(William Wimsatt)—他應該是最早指出,生物學家的工作不在於研究生物學是否可以被化約至物理與化學(也就是理論化約(theory reduction))的哲學家,溫薩特指出,生物學家實作的目標在於以化約的研究策略來發現機制(Wimsatt 1972: 67, 1976: 671, 1980),這部分我們會在第三節詳述。值得一提的是,溫薩特的新機制論立場與上述貝克托、理查森、格林蘭、麥契爾、達登、以及克瑞弗立場均有不同,雖然他主張以化約和建立機制模型的方法論來發現機制(Wimsatt 2002; 2007, ch. 6),但相較之下,他更看重生物現象的突現性質(Wimsatt 2000),也重視關於生物過程的形上學概念,如機敏性(heuristics)、強健性(robustness)、生成確立性(generative entrenchment)等等,這部分的詳細論述可見溫薩特2007年的著作《為有限的存有,重新建造哲學》(Re-Engineering Philosophy for Limited Beings)。除此之外,還有其他哲學家也有對機制提出刻畫,如泰伯瑞(Tabery 2004)、伊拉瑞和威廉森(Illari and Williamson 2012)、拉維(Levy 2013)、安德森(Andersen 2014a, 2014b)。
[4] 本段文字改寫自《生化學的原理》(Principle of biochemistry)「DNA複製」的段落。
[5] 關於理論與模型的中文科哲著作可見陳瑞麟於2012出版的《認知與評價:科學理論與實驗的動力學》之第一部分:模型與理論,特別是第一章到第四章。
[7] 科哲家蘇利文(Jacqueline Sullivan)、蘭吉諾(Helen Longino)、和泰伯瑞都曾為行為科學領域裡是否存有整合機制的可能性有過一番爭論(Sullivan 2009; Longino 2013, 2014; Tabery 2014a, 2014b)。
[8] 陳瑞麟對達登選用「策略」一詞的解析如下:「對達頓來說,策略是指引理論發展的程序(precedure)、實作(practice)、原則(principle),類似人工智能學家對於heuristics的用法,它是一個能產生合理結果的推理理方法(reasoning method),但對比於算則,算則保證產生正確的結果,啟迪則否。啟迪比算則更抽象,它可以說算是算則以上的指導原則,一個啟迪可能指導幾個不同的算則。然而達頓比較偏愛使用strategy這個詞,因為這個詞(比啟迪、算則)更一般…後來達頓直接使用『推理策略』(reasoning strategies)這個詞(Darden 2006)。」(陳瑞麟,《認知與評價》,頁557-558)
[9] 上節提過,格林蘭認為無論是機制框架或格架都是機制模型(mechanistic model),都表徵了欲發現機制的結構,在機制被完全發現前,都扮演了科學家用以推理與表徵的角色,所以其實就是模型。他以科學家發現人類說話知覺所提出來的競爭模型為例,論證從機制模型到完整機制描述是一段連續、不間斷的過程,由此質疑達登把機制區分為框架與格架等類,事實上,機制發現的歷程無法區分(Glennan 2005)。達登對此質疑的回應是:「模型」這個語詞太模糊,不易把發現機制所經歷的不同階段特徵呈現出來,實作上,這些階段確實為不同的階段(Darden 2006: 282; Darden 2007)。
[10] 克里克(Francis Crick)於1958年再次提出「中心法則」的概念,並在1970年《自然》(Nature)裡說道:「分子生物學的中心法則旨在詳細說明序列訊息的傳遞。它指明遺傳訊息不會從蛋白質傳遞至蛋白質或核苷酸。」(Crick 1970: 561-563)
[11] 關於對現象的刻畫,除了前面提到的觸發、調整、和抑制條件外,克瑞弗和達登也指出像「非標準條件」(nonstandard conditions)與「副作用」(by-products)(或「邊際效應」(side effect))等其他現象。非標準條件是指在實驗室以外的情況,例如在野外裡。副作用是指不具有「功能重要性」(functional signifance)的現象,但是雖說不具有功能重要性,但它對科學家在刻畫實驗現象時,還是會提供重要研究資訊(Craver and Darden 2013: 60)。
[12] 關於機制在演化論的討論包括Skipper and Millstein 2005; Baker 2005; Millstein 2006; Barros 2008; Hastad 2011; Mattewson and Calcott 2011; Illari and Williamson 2012; DesAutels 2018。在系統生物學有Bechtel 2006; Brigandt, Green, and O’Malley 2018。關於生態學的機制有Paslaru 2018。認知科學的機制討論有Thagard 2006; Bechtel 2008; Piccinini and Craver 2011; Weiskpf 2011; Stinson and Sullivan 2018。發育學則有Love 2018。在生醫領域有Clarke and Russo 2018。在經濟學有Marchionni 2018,以及神經經濟學(neuroeconomics,研究人類在作經濟決策時的生理機制)有Craver and Alexandrova 2008。行為遺傳學方面有Tabery 2014。
[13] 關於分子生物學中的機制,其實一直都有哲學家在進行反覆反省(Baetu 2018)。關於電腦計算機制請見Piccinini 2018; 工程科學方面見van Eck 2018。
[14] 摩根是國際知名的經濟思想史與經濟學哲學家,在科學哲學模型觀點上具有一席之地,是趙相科教授於荷蘭阿姆斯特丹大學博士論文的指導教授。
[15] 對此達登也有新回應(Darden 2015,中文翻譯見葉筱凡譯)。台灣中山大學萬毓澤(2015)也有相關討論,萬教授是從孔恩(Thomas Kuhn)的「演化轉向」切入談演化論和社會科學研究的關係。
一、中文文獻
洪純道,孫興祥(2000),〈海洋生物在神經科學的應用與貢獻〉,《國立海洋科技大學博物館籌備處簡訊》第9期,頁1-3。
陳瑞麟(2009),〈孟德爾究竟發現了什麼?一個實驗發現的典型模式〉,《科技、醫療與社會》第9期,頁123-172 。
陳瑞麟(2012),《認知與評價:科學理論與實驗的動力學》。台北:台大出版中心。
萬毓澤(2015),〈如何停止憂慮並愛上演化思維:從孔恩的「演化轉向」到「一般達爾文主義〉,《科技、醫療與社會》第20期,頁 181-222。
葉筱凡譯(2015),〈演化變遷的多層機制〉,《科技、醫療與社會》第21期,頁209-34。譯自Lindley Darden (2015). “Multilevel Mechanisms of Evolutionary Change.” Keynote Speaker Lecture.
趙相科(2019),〈經濟理論與模型〉,王一奇(編),《華文哲學百科》(2019版本)。
二、外文文獻
Allen, Garland E. (2018). “Mechanisms, Organicism, and Vitalism.” In Stuart Glennan and Phyllis Illari (eds.). The Routledge Handbook of Mechanisms and Mechanical Philosophy. London: Routledge.
Andersen, Holly (2012). “The Case for Regularity in Mechanistic Causal Explanation.” Synthese 189(3): 415-432.
Andersen, Holly (2014a). “A Field Guide to Mechanisms: Part I.” Philosophy Compass 4: 274–283.
Andersen, Holly (2014b). “A Field Guide to Mechanisms: Part II.” Philosophy Compass 4: 283–297.
Au, Yin-Chung (2019). “Time-Event Relationships as Representations for Constructing Cell Mechanisms.” In Samantha Kleinberg (ed.). Time and Causality Across the Sciences. Cambridge: Cambridge University Press.
Bacon, Francis [1620] 2000. The New Organon Cambridge: Cambridge University Press.
Bacon, Francis [1627] 2002. “The New Atlantis.” In Francis Bacon, the Major Works. Oxford: Oxford University Press, Oxford World’s Classics.
Baetu, Tudor (2018). “Mechanisms in Evolutionary Biology.” In Sturart Glennan and Phyllis Illari (eds.). The Routledge Handbook of Mechanisms and Mechanical Philosophy (pp. 308-318). London: Routledge.
Baker, J.M. (2005). “Adaptive Speciation: The Role of Natural Selection in Mechanisms of Geographic and Non-geographic Speciation.” In Craver and Darden 2005: 303–326.
Barros, D. Benjamin (2008). “Natural Selection as a Mechanism.” Philosophy of Science 75: 306-322.
Batterman, Robert W. and Collin C. Rice. (2014). “Minimal Model Explanations.” Philosophy of Science 81(3): 349-376.
Bechtel, William (2008). Mental Mechanisms: Philosophical Perspectives on Cognitive Neuroscience. London: Routledge.
Bechtel, William and Adele Abrhamsen (2005). “Explanation: A Mechanist Alternative.” In Carl F. Craver and Lindley Darden (eds.). Special Issue: “Mechanism in Biology.” Studies in History and Philosophy of Biological and Biomedical Science 36(2): 421-441.
Bechtel, William and Robert C. Richardson (2010 [1993]). Discovering Complexity: Decomposition and Localization as Strategies in Scientific Research. Second Edition. Cambridge, MA: MIT Press/Bradford Books.
Bogen, Jim (2005). “Regularities and Causality; Generalizations and Causal Explanations.” Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences 36 (2): 397-420.
Bogen, Jim (2008). “The New Mechanical Philosophy.” Metascience 17(1): 33-41.
Brigandt, Ingo, Sara Green, and Maureen A. O’Malley (2018). “Systems Biology and Mechanistic Explanation.” In Stuart Glennan and Phyllis Illari (eds.). The Routledge Handbook of Mechanisms and Mechanical Philosophy (pp. 362-374). London: Routledge.
Cartwright, Nancy (1989). Nature’s Capacity and Their Measurement. Oxford: Oxford University Press.
Cartwright, Nancy (2007). Hunting Causes and Using Them: Approaches in Philosophy and Economics. Cambridge: Cambridge University Press.
Chao, Hsiang-Ke and Szu-Ting Chen (2013). “Explaining the Explanations of 100 Million Missing Women.” In Hsiang-Ke Chao, Szu-Ting Chen, and L. Roberta Millstein (eds.) Mechanism and Causality in Biology and Economics. Dordrecht: Springer Netherlands.
Chen, Ruey-Lin (2013). “Experimental Discovery, Data Models, and Mechanisms in Biology.” In Hsiang-Ke Chao, Szu-Ting Chen, and L. Roberta Millstein (eds.) Mechanism and Causality in Biology and Economics. Dordrecht: Springer Netherlands.
Chen, Ruey-Lin (2017). “Mechanism, Capacities, and Nomological Machines.” In Hsiang-Ke Chao and Julian Reiss (eds.). Philosophy of Science in practice: Nancy Cartwright and the Nature of Scientific Reasoning (pp. 127-145). Synthesis Library, Vol. 379. Switzerland: Springer Press.
Clarke, Brendan and Federica Russo (2018). “Mechanisms and Biomedicine.” In Sturart Glennan and Phyllis Illari (eds.). The Routledge Handbook of Mechanisms and Mechanical Philosophy (pp. 319-331). London: Routledge.
Craver, Carl F. (2001). “Role Functions, Mechanisms and Hierarchy.” Philosophy of Science 68: 31–55.
Craver, Carl F. (2002). “Interlevel Experiments, Multilevel Mechanisms in the Neuroscience of Memory.” Philosophy of Science (Supplement) 69: S83-S97.
Craver, Carl F. (2006). “When Mechanistic Models Explain.” Syntheses 153(3): 355-376.
Craver, Carl F. (2007). Explaining the Brain: Mechanisms and the Mosaic Unity of Neuroscience. New York: Oxford University Press.
Craver, Carl F. (2012). “Functions and Mechanisms: A Perspectivalist View.” In Philippe Huneman (ed.) Functions: Selections and Mechanisms. Dordrecht: Springer.
Craver, Carl F. and Anna Alexandrova (2008). “No Revolution Necessary: Neural Mechanisms for Economics.” Economics and Philosophy 24: 381–406.
Craver, Carl F. and Lindley Darden (2001). “Discovering Mechanisms in Neurobiology: The Case of Spatial Memory.” In Peter Machamer, Rick Grush and Peter McLaughlin (eds.). Theory and Method in the Neuroscience. Pittsburgh, PA: University of Pittsburgh Press, pp. 112-137.
Craver, Carl F. and Lindley Darden (2013). In Search of Mechanisms: Discoveries Across the life Sciences. Chicago: University of Chicago Press.
Craver, Carl F. and James G. Tabery (2015). “Mechanisms in Science.” In Edward N. Zalta (ed.). The Stanford Encyclopedia of Philosophy, edited by Edward N. Zalta, Winter 2015. https://plato.stanford.edu/entries/science-mechanisms/
Crick, Francis (1970). Central Dogma of Molecular Biology. Nature 227(5258): 561-563.
Darden, Lindley (1991). Theory Change in Science: Strategies from Mendelian Genetics. New York: Oxford University Press.
Darden, Lindley (2005). “Relations Among Fields: Mendelian, Cytological and Molecular Mechanisms”, Studies in History and Philosophy of Biological and Biomedical Sciences, 36: 349–371.
Darden, Lindley (2006). Reasoning in Biological Discoveries: Essays on Mechanisms, Interfield Relations, and Anomaly Resolution. New York: Cambridge University Press.
Darden, Lindley (2007). "Mechanisms and Models.” In David L. Hull and Michael Ruse (eds.) Cambridge Companion to Philosophy of Biology. New York: Cambridge University Press.
Darden, Lindley (2008). “Thinking Again about Biological Mechanisms.” Philosophy of Science 75(5): 958-969.
Darden Lindley (2013). “Mechanisms Versus Causes in Biology and Medicine.” In Hsiang-Ke Chao, Szu-Ting Chen, and L. Roberta Millstein (eds.) Mechanism and Causality in Biology and Economics. Dordrecht: Springer Netherlands.
Darden, Lindley and Joseph Cain (1989.) “Selection Type Theories.” Philosophy of Science 56(1): 106–129.
Darden, Lindley and Carl F. Craver (2002). “Strategies in the Interfield Discovery of the Mechanism of Protein Synthesis.” Studies in History and Philosophy of Biology and Biomedical Sciences 33: 1-28. Corrected and reprinted in Darden 2006, ch. 3.
DesAutels, Lane (2018). “Mechanisms in Evolutionary Biology.” In Sturart Glennan and Phyllis Illari (eds.). The Routledge Handbook of Mechanisms and Mechanical Philosophy (pp. 296-307). London: Routledge.
Giere, Ronald N. (1988). Explaining Science: A Cognitive Approach. Chicago: University of Chicago Press.
Giere, Ronald N. (1999). “Using Models to Represent Reality.” In L. Magnani, N. Nersessian, and P. Thagard (eds.). Model-Based Reasoning in Scientific Discovery. New York: Springer, pp. 41-57.
Giere, Ronald N. (2004). “How Models Are Used to Represent Reality.” Philosophy of Science 71(5): 742-752.
Giere, Ronald N. (2006). Scientific Perspectivism. Chicago: University of Chicago Press.
Glennan, Stuart S. (1996). “Mechanisms and the Nature of Causation.” Erkenntnis 44(1): 49–71.
Glennan, Stuart S. (2002). “Rethinking Mechanistic Explanation.” Philosophy of Science (Supplement) 69: S342-S353.
Glennan, Stuart S. (2005). “Modeling Mechanisms.” Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences 36(2): 443-464.
Glennan, Stuart S. (2017). The New Mechanical Philosophy. Oxford: Oxford University Press.
Havstad, Joyce C. (2011). “Discussion: Problems for Natural Selection as a Mechanism”. Philosophy of Science 78: 512–523.
Hempel, Carl G. (1965). Aspects of Scientific Explanation and Other Essays in the Philosophy of Science. New York: Free Press.
Hemple, Carl G. and Paul Oppenheim (1948). “Studies in the Logic of Explanation.” Philosophy of Science 15(2): 135-175.
Hull, David (1974). Philosophy of Biological Science. Englewood Cliffs, NJ: Prentice-Hall.
Illari, Phyllis and Jon Williamson (2012). “What is a Mechanism?: Thinking about Mechanisms Across the Sciences.” European Journal for Philosophy of Science 2: 119–135.
Kaplan, David Michael and William Bechtel (2011). “Dynamical Models: An Alternative or Complement to Mechanistic Explanations?” Topics in Cognitive Science 3(2): 438-444.
Kaplan, David Michael, and Carl F. Craver (2011). “The Explanatory Force of Dynamical and Mathematical Models in Neuroscience: A Mechanistic Perspective.” Philosophy of Science 78(4): 601-627.
Kitcher, Phillip (1984). 1953 and all that: A Tale of Two Science. Philosophical Review 93: 335-373.
Kirckel, Beate (2014). The Metaphysics of Mechanism. Humboldt-Universität zu Berlin.
Lakatos, Imre (1978). The Methodology of Scientific Research Programmes: Philosophical Papers, vol. 1. Cambridge: Cambridge University Press.
Levy, Arnon (2013). “Three Kinds of New Mechanism.” Biology and Philosophy 28: 99-114.
Levy, Arnon and William Bechtel (2013). “Abstraction and the Organization of Mechanisms.” Philosophy of Science 80(2): 241-261.
Longino, Helen (2013). Studying Human Behavior: How Scientists Investigate Aggression and Sexuality. Chicago: University of Chicago Press.
Longino, Helen (2014). “Pluralism, Social Action, and the Causal Space of Human Behavior.” Metascience 23: 443–459.
Love, Alan (2018). “Develomental Mechanisms.” In Sturart Glennan and Phyllis Illari (eds.). The Routledge Handbook of Mechanisms and Mechanical Philosophy (pp. 332-347). London: Routledge.
Leuridan, Bert (2010). “Can Mechanisms Really Replace Laws of Nature.” Philosophy of Science 77: 317-340.
Machamer, Peter, Lindley Darden, and Carl F. Craver (2000). “Thinking about Mechanisms.” Philosophy of Science 67(1): 1–25.
Machamer, Peter (2004). “Activities and Causation: The Metaphysics and Epistemology of Mechanisms.” International Studies in the Philosophy of Science 18(1): 27-39.
Matthewson, John and Brett Calcott (2011). “Mechanistic Models of Population-Level Phenomena.” Biology and Philosophy 26: 737–756.
Marchionni, Caterina (2018). “Mechanisms in Economics.” In Sturart Glennan and Phyllis Illari (eds.). The Routledge Handbook of Mechanisms and Mechanical Philosophy (pp. 423-434). London: Routledge.
Millstein, Roberta L. (2006). “Natural Selection as a Population-Level Causal Process.” The British Journal for the Philosophy of Science 57: 627 -653.
Mitchell, Sandra (1997). “Pragmatic Laws.” Philosophy of Science (Supplement) 64: S468-S479.
Nersessain, Nancy I. (1992). How Do Scientists Think? Capturing the Dynamics of Conceptual Change in Science. In Ronald Giere (ed.). Cognitive Models of Science (pp. 3-44). Minneapolis: University of Minnesota Press.
Nagel, Ernest (1961). The Structure of Science: Problems in the Logic of Scientific Explanation. New York: Harcourt, Brace and World.
Paslaru, Viorel (2018). “Mechanisms in Ecology.” In Sturart Glennan and Phyllis Illari (eds.). The Routledge Handbook of Mechanisms and Mechanical Philosophy (pp. 348-361). London: Routledge.
Piccinini, Gualtiero and Carl F. Craver (2011). “Integrating Psychology and Neuroscience: Functional Analyses as Mechanism Sketches.” Synthese 183: 283–311.
Piccinini, Gualtiero (2018). “Computational Mechanisms.” In Sturart Glennan and Phyllis Illari (eds.). The Routledge Handbook of Mechanisms and Mechanical Philosophy (pp. 435-446). London: Routledge.
Popper, Karl (1959). The Logic of Scientific Discovery. London: Hutchinson and Co.
Rheinberger, Hans-Jörg (1997). Towers a History of Epistemic Things: Synthesizing Protein in the Test Tube. Palo Alto, CA: Stanford University Press.
Rosenberg, Alexander (1985). The Structure of Biological Science. Cambridge, UK: Cambridge University Press.
Schaffner, Kenneth (1974). “Logic of Discovery and Justification in Regulatory Genetics.” Studies in History and Philosophy of Science 4: 349-385.
Schaffner, Kenneth (1993). Discovery and Explanation in Biology and Medicine. Chicago, IL: University of Chicago Press.
Stinson, Catherine and Jacqueline Sullivan (2018). “Mechanisms in Evolutionary Biology.” In Sturart Glennan and Phyllis Illari (eds.). The Routledge Handbook of Mechanisms and Mechanical Philosophy (pp. 375-388). London: Routledge.
Skipper, Robert A. Jr., and Roberta L. Millstein (2005). “Thinking about Evolutionary Mechanisms: Natural Selection.” Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences 36 (2): 327-347.
Steel, Daniel (2008). Across the Boundaries: Extrapolation in Biology and Social Science. New York: Oxford University Press.
Strevens, Michael (2008). Depth: An Account of Scientific Explanation. Cambridge, MA: Harvard University Press.
Sullivan, Jacqueline (2009). “The Multiplicity of Experimental Protocols: A Challenge to Reductionist and Non-Reductionist Models of the Unity of Neuroscience.” Synthese 167: 511–539.
Suppe, Frederick (1997). The Structure of Scientific Theories. Chicago: University of Illinois Press.
Tabery, James (2004). “Synthesizing Activities and Interactions in the Concept of a Mechanism.” Philosophy of Science 71: 1-15.
Tabery, James (2014a). Beyond Versus: The Struggle to Understand the Interaction of Nature and Nurture. Cambridge, MA: The MIT Press.
Tabery, James (2014b). “Pluralism, Social Action, and the Causal Space of Human Behavior.” Metascience 23: 443–459.
Thagard, Paul (2006). Hot Thought: Mechanisms and Applications of Emotional Cognition. Cambridge, MA: The MIT Press.
van Eck, Dingmar (2018). “Mechanisms and Engineering Science.” In Sturart Glennan and Phyllis Illari (eds.). The Routledge Handbook of Mechanisms and Mechanical Philosophy (pp. 447-461). London: Routledge.
van Fraassen, Bas C. (1980). The Scientific Image. Oxford: Oxford University Press.
Wong, Rong-Lin (2013). “Is Natural Selection a Population-Level Causal Process?” In Hsiang-Ke Chao, Szu-Ting Chen, and L. Roberta Millstein (eds.) Mechanism and Causality in Biology and Economics. Dordrecht: Springer Netherlands.
Weiskopf, Daniel (2011). “Models and Mechanisms in Psychological Explanation.” Synthese 183: 313–338.
Wimsatt, William (1972). “Complexity and Organization.” In PSA: Proceedings of the Biennial Meeting of the Philosophy of Science Association Vol. 1972, edited by Kenneth Schaffner and Robert Cohen, 67-86. Dordrecht: D. Reidel.
Wimsatt, William (1976). “Reductionism, Levels of Organization, and the Mind–Body Problem.” In G. Globus, I. Savodnik, and G. Maxwell (eds), Consciousness and the Brain, New York: Plenum Press, pp. 199–267.
Wimsatt, William (1980). “Reductionistic Research Strategies and Their Biases in the Units of Selection Controversy.” In Thomas Nickles (ed.), Scientific Discovery: Case Studies, Dordrecht: Reidel, pp. 213-259.
Wimsatt, William (2000). “Emergence as Non-Aggregativity and the Biases of Reductionisms.” Foundations of Science 5(3): 269-197.
Wimsatt, William (2002). “Using False Models to Elaborate Constraint on Process: Blending Inheritance in Organic and Cultural Evolution.” Philosophy of Science 69(S3): S12-24.
Wimsatt, William (2007). Re-Engineering Philosophy for Limited Beings. Cambridge, MA: Harvard University Press.
Woodward, James (2000). “What Is a Mechanism? A Counterfactual Account.” Philosophy of Science 69(S3): S366-377.
Woodward, James (2003). Making Things Happen. Oxford: Oxford University Press.
Woodward, James (2014). “Simplicity in the Best Systems Account of Laws of Nature.” British Journal for Philosophy of Science 65: 91-123.
Yeh, Hsiao-Fan, & Chem, Ruey-Lin (2017). An Experiment-Based Methodology for Classical Genetics and Molecular Biology. Annals of the Japan Association for Philosophy of Science, 26, 39-60.